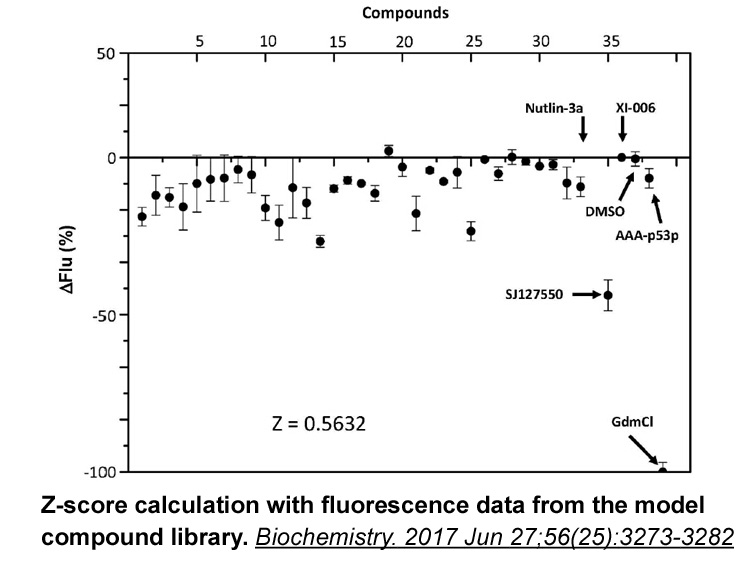
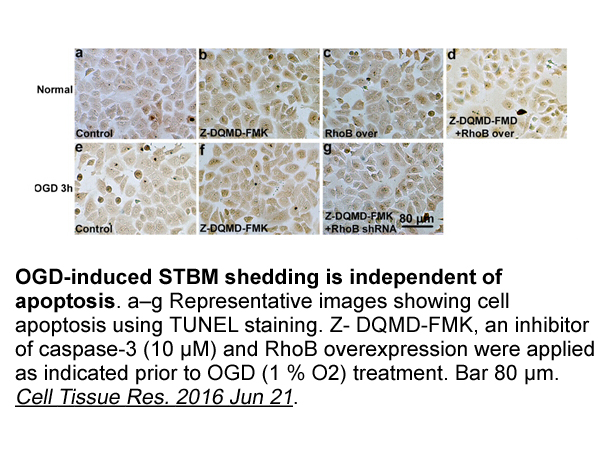
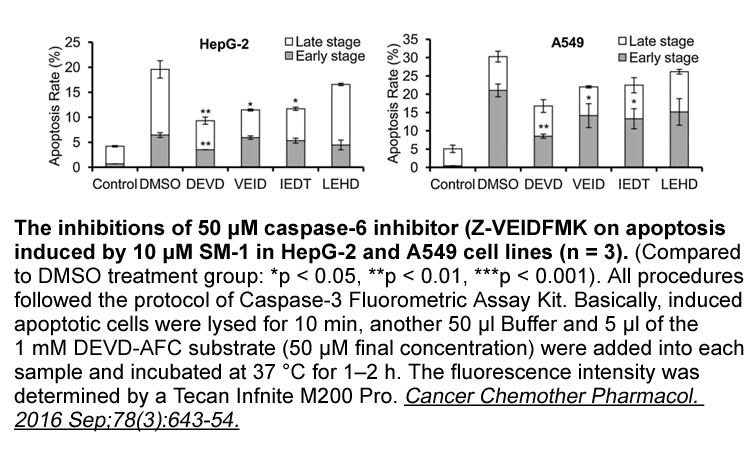
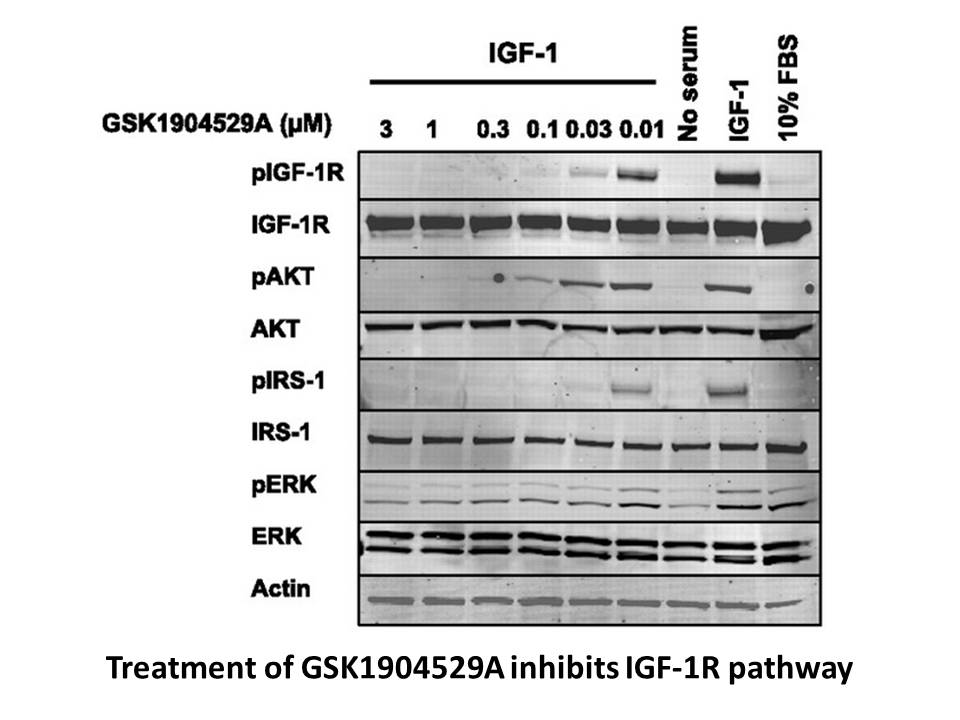
Archives
- 2026-04
- 2026-03
- 2026-02
- 2026-01
- 2025-12
- 2025-11
- 2025-10
- 2025-09
- 2025-03
- 2025-02
- 2025-01
- 2024-12
- 2024-11
- 2024-10
- 2024-09
- 2024-08
- 2024-07
- 2024-06
- 2024-05
- 2024-04
- 2024-03
- 2024-02
- 2024-01
- 2023-12
- 2023-11
- 2023-10
- 2023-09
- 2023-08
- 2023-07
- 2023-06
- 2023-05
- 2023-04
- 2023-03
- 2023-02
- 2023-01
- 2022-12
- 2022-11
- 2022-10
- 2022-09
- 2022-08
- 2022-07
- 2022-06
- 2022-05
- 2022-04
- 2022-03
- 2022-02
- 2022-01
- 2021-12
- 2021-11
- 2021-10
- 2021-09
- 2021-08
- 2021-07
- 2021-06
- 2021-05
- 2021-04
- 2021-03
- 2021-02
- 2021-01
- 2020-12
- 2020-11
- 2020-10
- 2020-09
- 2020-08
- 2020-07
- 2020-06
- 2020-05
- 2020-04
- 2020-03
- 2020-02
- 2020-01
- 2019-12
- 2019-11
- 2019-10
- 2019-09
- 2019-08
- 2019-07
- 2019-06
- 2019-05
- 2019-04
- 2018-11
- 2018-10
- 2018-07
-
Intriguingly one E residue may serve as a molecular gate
2019-12-06
Intriguingly, one E2 residue may serve as a molecular ‘gate’ to allow the C-terminus of ubiquitin to access the closed E2~Ub conformations favorable for ubiquitin transfer. This residue, Asp87 in UbcH5 family members, resides on one side of the opening that leads to the active site Cys (Fig. 4D). In
-
The murine subfamily members including and have
2019-12-06

The murine subfamily members including , , , and have been identified and their encoded proteins showed deubiquitinating enzymatic activity , , , . It has been demonstrated that is an immediate-early gene induced by IL-3 in B-lymphocytes and is an IL-2 specific immediate-early gene in T-lymphoc
-
Mammalian cells have two distinct DNA repair pathways for
2019-12-06
Mammalian CC-4047 have two distinct DNA repair pathways for DSBs, NHEJ and HR. Our data are consistent with the suggestion that the DNA–PK-mediated NHEJ pathway recognizes DSBs faster than the HR pathway and acts before the activation of the DNA damage S-phase checkpoint. The activation of NHEJ by
-
After incorporation of random nucleotides by
2019-12-06
After incorporation of random histone demethylase inhibitor by TdT during heavy-chain rearrangements, both TdT and pol λ may perform in trans polymerase activity (in unknown proportions), whereas synthesis of the complementary strand can only be achieved by pol λ using its gap-filling activity, whic
-
The absence of the LIG
2019-12-06

The absence of the LIG3 gene in yeasts has prevented the use of genetically tractable lower eukaryotes, such as Saccharomyces cerevisiae and Schizosaccharomyces pombe, as models to gain insights into the cellular functions of and interplay between the DNA ligases encoded by the LIG1, LIG3 and LIG4 g
-
br Conclusion br Competing interests br Authors contribution
2019-12-06

Conclusion Competing interests Authors\' contributions Acknowledgements The work was supported by Aurigene Discovery Technologies (M) Sdn. Bhd., Industrial Doctoral Program sponsorship by Ministry of Higher Education, Malaysia and research grants by the University of Malaya, grant number
-
Substituents at the C position An oxygen containing substitu
2019-12-06
Substituents at the C11 position — An oxygen-containing substituent at the C11 position also affects the catalytic activity of Δ1-KSTDs. The Δ1-KSTDs from the Gram-positive bacteria R. equi [29], N. simplex ATCC 6946 and IFO 12069 [48,49,52], and R. rhodochrous IFO 3338 [27] were able to 1(2)-dehydr
-
br Cysteine proteases as molecular
2019-12-06
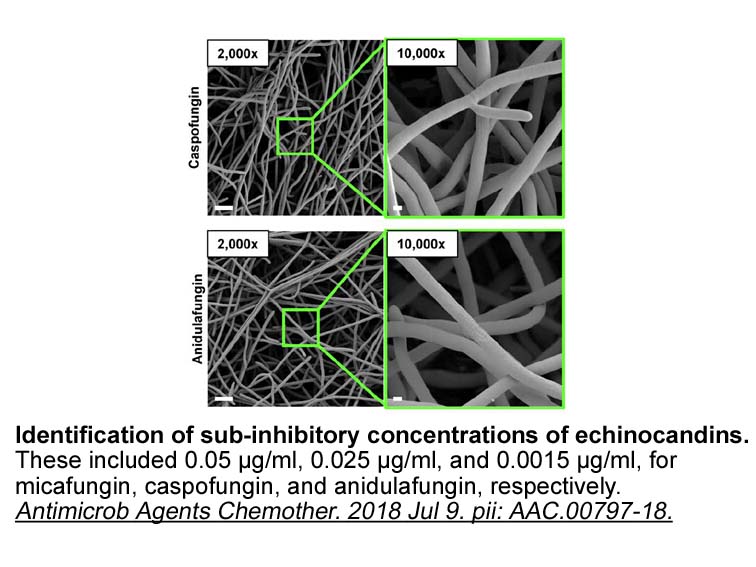
Cysteine proteases as molecular targets for trypanosomatid diseases Approaches relying on a molecular target, such as SBDD and target-based screening, have supported outstanding scientific developments in current pharmaceutical R&D. By integrating experimental and computational methods, these str
-
br Conflicts of interest br Acknowledgments
2019-12-05

Conflicts of interest Acknowledgments This research was supported by the Basic Science Research Program through the National Research Foundation of Korea funded by the Ministry of Science, ICT and Future Planning (NRF-2015R1A2A2A01004593). Introduction Resistance to Inhibitors of Cholinest
-
br Acknowledgments This study was supported by grants from t
2019-12-05
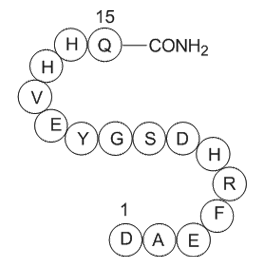
Acknowledgments This study was supported by grants from the Spanish Ministry of Economy and CompetitivenessPI10/00387, PI12/01087, PI12/01703, IPT-2011-0817-010000, and RIC Red de Investigación Cardiovascular (RIC)RD12/0042/0055. RIC is an initiative of Instituto de Salud Carlos III (ISCIII), Spa
-
br Introduction Concrete is one of
2019-12-05

Introduction Concrete is one of the most commonly used materials in the construction industry and its raw material is available throughout the world. Concrete is derived from ordinary Portland cement and is used for the construction and development of infrastructures. Research shows that demand f
-
NU 7026 At baseline mutant allele frequency of EGFR in tissu
2019-12-05
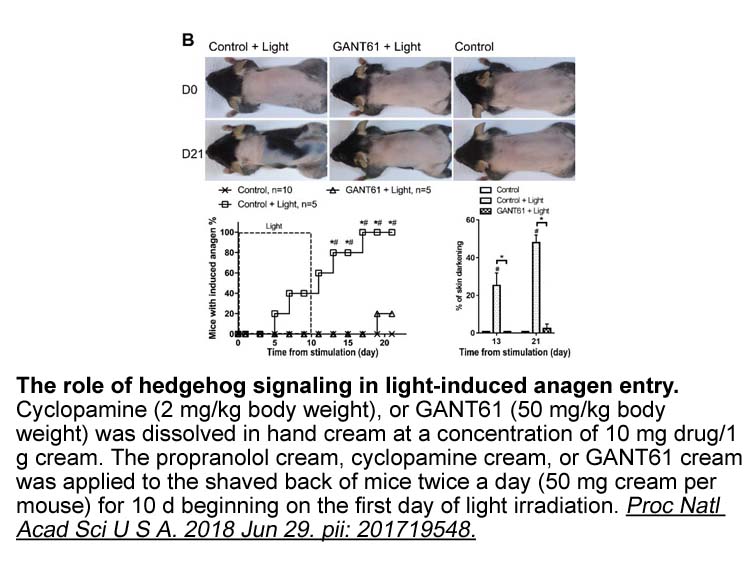
At baseline, mutant allele frequency of EGFR in tissue and plasma samples did not correlated with anti-tumor response (Supplementary Fig. 1). Among 35 patients who were positive for cell-free DNA (cfDNA) at baseline, changes of cfDNA during treatment were analyzed. Negative conversion (NC) of cfDNA
-
The biological significance of DNA ligases is attributed to
2019-12-04
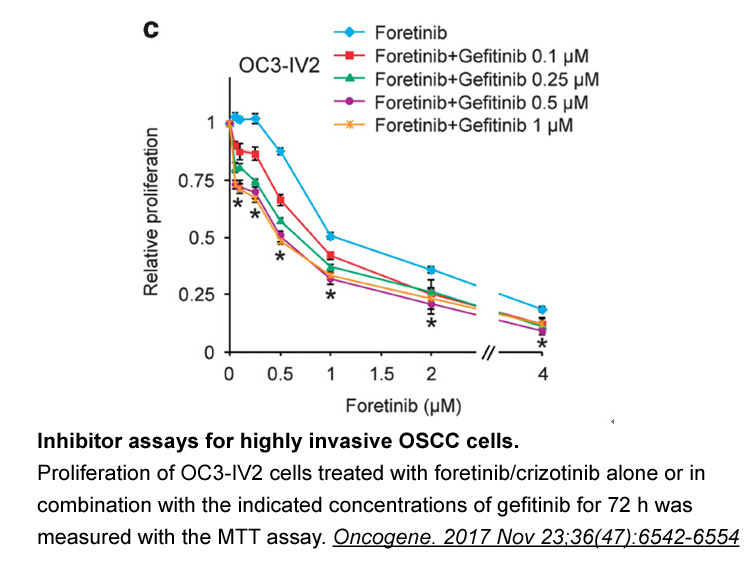
The biological significance of DNA ligases is attributed to their function in DNA replication, recombination and/or DNA repair [1], [2]. In vertebrates, three different DNA ligases I, III and IV are recognized. DNA ligase I is essential for the Okazaki fragments’ ligation during lagging-strand DNA s
-
LPA is known to regulate the peroxisome proliferator activat
2019-12-04
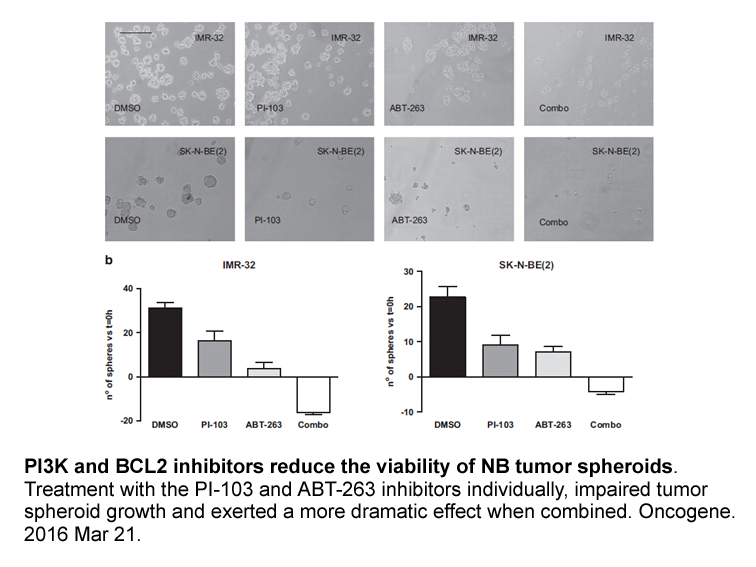
LPA is known to regulate the peroxisome proliferator-activated receptor γ and the reorganization of actin cytoskeleton [58]. However, the physiological functions of LPA in cells are still poorly understood. The simultaneous production of PA with/without 1-LPA or 2-LPA may be important to maximize a
-
Tadalafil Introduction The Discoidin Domain Receptors DDRs c
2019-12-04
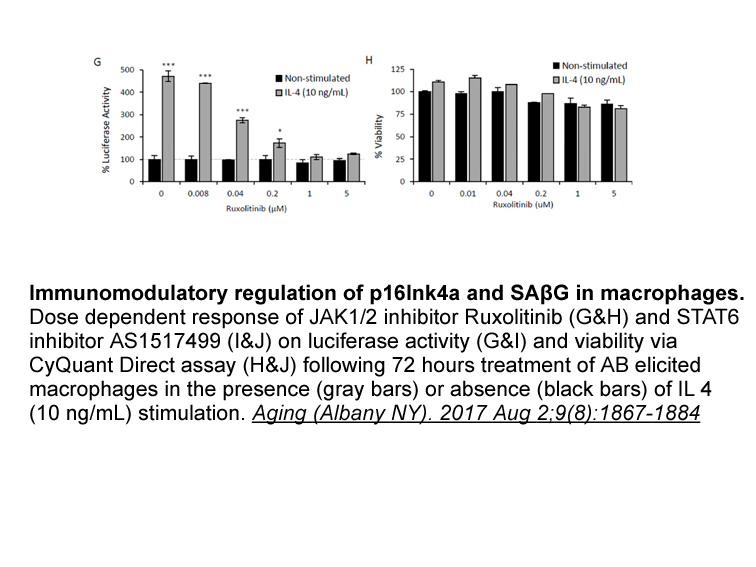
Introduction The Discoidin Domain Receptors (DDRs), comprising DDR1 and DDR2, are collagen-binding receptor tyrosine kinases (RTKs) that function as microenvironmental sensors at the interface of the extracellular matrix and the intracellular signal transduction machinery [1]. In response to ligand
16191 records 925/1080 page Previous Next First page 上5页 921922923924925 下5页 Last page